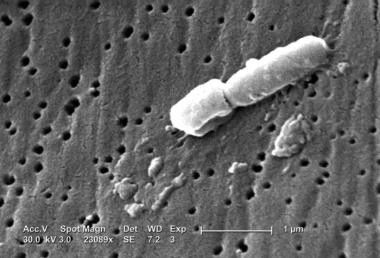
Proteus
Proteus mirabilis
Proteus vulgaris
Proteus Syndrome
Proteus penneri
Urease
Arachidonate Lipoxygenases
Benzeneacetamides
Tablets
Methenamine
4,5-Dihydro-1-(3-(trifluoromethyl)phenyl)-1H-pyrazol-3-amine
Identification of protease and rpoN-associated genes of uropathogenic Proteus mirabilis by negative selection in a mouse model of ascending urinary tract infection. (1/238)
Proteus mirabilis, a motile gram-negative bacterium, is a principal cause of urinary tract infections in patients with functional or anatomical abnormalities of the urinary tract or those with urinary catheters in place. Thus far, virulence factors including urease, flagella, haemolysin, various fimbriae, IgA protease and a deaminase have been characterized based on the phenotypic traits conferred by these proteins. In this study, an attempt was made to identify new virulence genes of P. mirabilis that may not have identifiable phenotypes using the recently described technique of signature-tagged mutagenesis. A pool of chromosomal transposon mutants was made through conjugation and kanamycin/tetracycline selection; random insertion was confirmed by Southern blotting of chromosomal DNA isolated from 16 mutants using the aphA gene as a probe. From the total pool, 2.3% (9/397) auxotrophic mutants and 3.5% (14/397) swarming mutants were identified by screening on minimal salts agar and Luria agar plates, respectively. Thirty per cent of the mutants, found to have either no tag or an unamplifiable tag, were removed from the input pool. Then 10(7) c.f.u. from a 96-mutant pool (approximately 10(5) c.f.u. of each mutant) were used as an input pool to transurethrally inoculate seven CBA mice. After 2 d infection, bacteria were recovered from the bladders and kidneys and yielded about 10(5) c.f.u. as an output pool. Dot blot analysis showed that two of the 96 mutants, designated B2 and B5, could not be hybridized by signature tags amplified from the bladder output pool. Interrupted genes from these two mutants were cloned and sequenced. The interrupted gene in B2 predicts a polypeptide of 37.3 kDa that shares amino acid similarity with a putative protease or collagenase precursor. The gene in B5 predicts a polypeptide of 32.6 kDa that is very similar to that encoded by ORF284 of the rpoN operon controlling expression of nitrogen-regulated genes from several bacterial species. The virulence of the two mutants was tested further by co-challenging CBA mice with each mutant and the parental strain. After 1 week of infection, the B2 and B5 mutants were recovered in numbers 100-fold and 1000-fold less than the parental strain, respectively. Using an in vitro assay, it was shown that the B2 mutant had significantly less (P = 0.0001) extracellular protease activity than the wild-type strain. These findings demonstrate that signature-tagged mutagenesis is a viable approach to identify bacterial genes associated with the ability to infect the urinary tract. (+info)Requirement of MrpH for mannose-resistant Proteus-like fimbria-mediated hemagglutination by Proteus mirabilis. (2/238)
Two new genes, mrpH and mrpJ, were identified downstream of mrpG in the mrp gene cluster encoding mannose-resistant Proteus-like (MR/P) fimbriae of uropathogenic Proteus mirabilis. Since the predicted MrpH has 30% amino acid sequence identity to PapG, the Galalpha(1-4)Gal-binding adhesin of Escherichia coli P fimbriae, we hypothesized that mrpH encodes the functional MR/P hemagglutinin. MR/P fimbriae, expressed in E. coli DH5alpha, conferred on bacteria both the ability to cause mannose-resistant hemagglutination and the ability to aggregate to form pellicles on the broth surface. Both a DeltamrpH mutant expressed in E. coli DH5alpha and an isogenic mrpH::aphA mutant of P. mirabilis were unable to produce normal MR/P fimbriae efficiently, suggesting that MrpH was involved in fimbrial assembly. Amino acid residue substitution of the N-terminal cysteine residues (C66S and C128S) of MrpH abolished the receptor-binding activity (hemagglutinating ability) of MrpH but allowed normal fimbrial assembly, supporting the notion that MrpH was the functional MR/P hemagglutinin. Immunogold electron microscopy of P. mirabilis HI4320 revealed that MrpH was located at the tip of MR/P fimbriae, also consistent with its role in receptor binding. The isogenic mrpH::aphA mutant of HI4320 was less able to colonize the urine, bladder, and kidneys in a mouse model of ascending urinary tract infection (P < 0.01), and therefore MR/P fimbriae contribute significantly to bacterial colonization in mice. While there are similarities between P. mirabilis MR/P and E. coli P fimbriae, there are more notable differences: (i) synthesis of the MrpH adhesin is required to initiate fimbrial assembly, (ii) MR/P fimbriae confer an aggregation phenotype, (iii) site-directed mutation of specific residues can abolish receptor binding but allows fimbrial assembly, and (iv) mutation of the adhesin gene abolishes virulence in a mouse model of ascending urinary tract infection. (+info)Growth, cellular differentiation and virulence factor expression by Proteus mirabilis in vitro and in vivo. (3/238)
A uropathogenic strain of Proteus mirabilis was grown in vitro in human and mouse urine and brain-heart infusion broth (BHIB) and in vivo in subcutaneous open chambers (SOC) in mice, intraperitoneal diffusion chambers (IPC) in rats and by ascending urinary tract infection in mice in order to compare growth pattern, cellular differentiation and expression of virulence factors. Although the growth rate was slower in vivo than in vitro, the extent of growth was similar after 24 h. PR mirabilis differentiated into filamentous swarmer cells in all in-vitro culture conditions, but no filamentous cells were observed in either of the in-vivo chamber models. Transurethrally infected mice showed a rapid release or loss of filamentous cells and these could not be seen in kidney or bladder homogenates 7 days after infection. Bacteria showed increasing haemagglutination titres for fresh and tanned red blood cells after subculturing in BHIB, but bacteria grown in vivo did not show haemagglutination. An increasing resistance to normal serum was found when bacteria were grown in vivo. Significant haemolytic activity was detected with bacteria grown in BHIB and IPC, but almost no activity was found when bacteria had grown in urine. These findings improve the understanding of the role of P. mirabilis uropathogenic virulence factors in vivo. (+info)Investigation of the types and characteristics of the proteolytic enzymes formed by diverse strains of Proteus species. (4/238)
Many diverse clinical isolates of Proteus mirabilis (48 strains), P. penneri (25), P. vulgaris biogroup 2 (48) and P. vulgaris biogroup 3 (21) from man were examined for their ability to produce proteolytic enzymes and the nature and characteristics of the proteases were studied. All the P. penneri isolates, most (94-90%) of the P. mirabilis and P. vulgaris biogroup 2 isolates, but only 71% of the P. vulgaris biogroup 3 isolates, secreted proteolytic enzymes. These were detected most readily at pH 8 with gelatin as substrate. A strong correlation was found between the ability of a strain to form swarming growth and its ability to secrete proteases. Non-swarming isolates invariably appeared to be non-proteolytic. However, some isolates, particularly of P. vulgaris biogroup 3, were non-proteolytic even when they formed swarming growth. Analysis of the secreted enzymes of the different Proteus spp. on polyacrylamide-gelatin gels under various constraints of pH and other factors showed that they were all EDTA-sensitive metalloproteinases. Analysis of the kinetics of production of the proteases revealed the formation of an additional protease of undefined type and function that was cell-associated and formed before the others were secreted. The secreted protease was subsequently modified to two isoforms whose mass (53-46 kDa) varied with the Proteus spp. and the strain. There was no evidence that the secreted proteases of strains of Proteus spp. were of types other than metalloproteinases. (+info)Structures of the O-specific polysaccharides and a serological cross-reactivity of the lipopolysaccharides of Proteus mirabilis O24 and O29. (5/238)
Strains of Proteus mirabilis belonging to serogroups O24 and O29 are frequent in clinical specimens. Anti-P. mirabilis O24 serum cross-reacted with the lipopolysaccharide (LPS) of P. mirabilis O29 and vice versa. The structures of the O-specific polysaccharides (OPSs, O-antigens) of both LPSs were established using sugar analysis and one- and two-dimensional 1H- and 13C-NMR spectroscopy and found to be different. SDS-PAGE and Western immunoblotting suggested that the serological cross-reactivity of the LPSs is due to a common epitope(s) on the core-lipid A moiety, rather than on the OPS. Therefore, the epitope specificity and the structures of the O-antigens studied are unique among Proteus serogroups. (+info)Serum immunoglobulin response and protection from homologous challenge by Proteus mirabilis in a mouse model of ascending urinary tract infection. (6/238)
We tested the hypothesis that experimental Proteus mirabilis urinary tract infection in mice would protect against homologous bladder rechallenge. Despite production of serum immunoglobulin G (IgG) and IgM (median titers of 1:320 and 1:80, respectively), vaccinated (infected and antibiotic-cured) mice did not show a decrease in mortality upon rechallenge; the survivors experienced only modest protection from infection (mean log(10) number of CFU of P. mirabilis Nal(r) HI4320 per milliliter or gram in vaccinated mice versus sham-vaccinated mice: urine, 100-fold less [3.5 versus 5.5; P = 0.13]; bladder, 100-fold less [3.1 versus 5.1; P = 0.066]; kidneys, 40-fold less [2.7 versus 4.3; P = 0.016]). Western blots using protein from the wild-type strain and isogenic mutants demonstrated antibody responses to MR/P and PMF fimbriae and flagella. There was no correlation between serum IgG or IgM levels and protection from mortality or infection. There was a trend toward elevated serum IgA titers and protection from subsequent challenge (P >/= 0.09), although only a few mice developed significant serum IgA levels. We conclude that prior infection with P. mirabilis does not protect significantly against homologous challenge. (+info)Problems related to determination of MICs of oximino-type expanded-spectrum cephems for Proteus vulgaris. (7/238)
During in vitro susceptibility testing of clinical isolates of Proteus vulgaris, we noted that the MICs of several expanded-spectrum cephems were much higher in the broth microdilution method than in the agar dilution method (termed the MIC gap phenomenon). Here we investigated the mechanism of the MIC gap phenomenon. Cephems with the MIC gap phenomenon were of the oximino type, such as cefotaxime, cefteram, and cefpodoxime, which serve as good substrates for inducible class A beta-lactamase (CumA) enzymes produced by P. vulgaris; this finding suggests a relationship between the MIC gap phenomenon and CumA. Since peptidoglycan recycling shares a system common to that inducing CumA, we analyzed the mechanism of the MIC gap phenomenon using P. vulgaris B317 and isogenic mutants with mutations in the peptidoglycan recycling and beta-lactamase induction systems. The MIC gap phenomenon was observed in the parent strain B317 but not in B317G (cumG-defective mutant; defective peptidoglycan recycling) and B317R (cumR-defective mutant; defective CumA transcriptional regulator). No beta-lactamase activity was detected in B317G and B317R. beta-Lactamase activity and the MIC gap phenomenon were restored in B317G/pMD301 (strain transcomplemented by a cloned cumG gene) and B317R/pMD501 (strain transcomplemented by a cloned cumR gene). MICs determined by the agar dilution method increased when lower agar concentrations were used. Our results indicated that the mechanism of the MIC gap phenomenon is related to peptidoglycan recycling and CumA induction systems. However, it remains unclear how beta-lactamase induction of P. vulgaris is suppressed on agar plates. (+info)Evaluation of the discriminatory powers of the Dienes test and ribotyping as typing methods for Proteus mirabilis. (8/238)
A total of 63 clinical isolates of Proteus mirabilis collected over a 19-month period were typed by the Dienes test and ribotyping. Ribotyping was performed using the fully automated RiboPrinter Microbial Characterization System (Qualicon, Wilmington, Del.). Isolates that were indistinguishable by the Dienes test and/or ribotyping were characterized further by pulsed-field gel electrophoresis (PFGE). Most of the isolates represented unique strains as judged by the Dienes test and ribotyping. Forty isolates represented 40 different ribotypes and Dienes types. The remaining 23 isolates were grouped into 13 Dienes types, 12 ribotypes, and 14 PFGE types. The index of discrimination was 0.980 for the Dienes test, 0.979 for ribotyping, and 0.992 for PFGE. Both the Dienes test and ribotyping are useful methods for identifying individual strains of P. mirabilis. The Dienes test is simple, inexpensive, and easy to perform. It can be performed in virtually any laboratory and should be used in the initial epidemiologic characterization of P. mirabilis isolates. (+info)Proteus infections are caused by the bacterium Proteus mirabilis or other Proteus species. These bacteria are gram-negative, opportunistic pathogens that can cause various types of infections, including urinary tract infections (UTIs), wound infections, and bacteremia (bloodstream infections). Proteus infections are often associated with complicated UTIs, catheter-associated UTIs, and healthcare-associated infections. They can be difficult to treat due to their ability to produce enzymes that inactivate certain antibiotics and form biofilms.
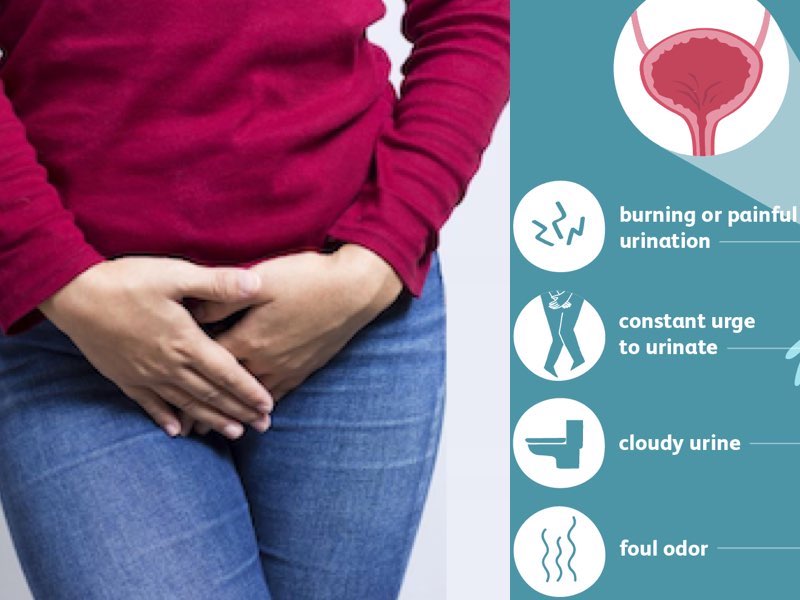
Proteus infections can cause symptoms such as fever, chills, fatigue, and discomfort in the affected area. In UTIs, patients may experience symptoms like burning during urination, frequent urges to urinate, and cloudy or foul-smelling urine. Wound infections caused by Proteus can lead to delayed healing, increased pain, and pus formation. Bacteremia can cause sepsis, a life-threatening condition that requires immediate medical attention.
Treatment for Proteus infections typically involves antibiotics, such as fluoroquinolones, trimethoprim-sulfamethoxazole, or carbapenems. The choice of antibiotic depends on the severity and location of the infection, as well as the patient's overall health status and any underlying medical conditions. In some cases, surgical intervention may be necessary to drain abscesses or remove infected devices like catheters.
'Proteus' doesn't have a specific medical definition itself, but it is related to a syndrome in medicine. Proteus syndrome is a rare genetic disorder characterized by the overgrowth of various tissues and organs in the body. The name "Proteus" comes from the Greek god Proteus, who could change his form at will, reflecting the diverse and ever-changing nature of this condition's symptoms.
People with Proteus syndrome experience asymmetric overgrowth of bones, skin, and other tissues, leading to abnormalities in body shape and function. The disorder can also affect blood vessels, causing benign tumors called hamartomas to develop. Additionally, individuals with Proteus syndrome are at an increased risk of developing certain types of cancer.
The genetic mutation responsible for Proteus syndrome is found in the AKT1 gene, which plays a crucial role in cell growth and division. This disorder is typically not inherited but instead arises spontaneously as a new mutation in the affected individual. Early diagnosis and management of Proteus syndrome can help improve patients' quality of life and reduce complications associated with the condition.
Proteus mirabilis is a species of Gram-negative, facultatively anaerobic, rod-shaped bacteria that are commonly found in the environment, particularly in soil and water. In humans, P. mirabilis can be part of the normal gut flora but can also cause opportunistic infections, particularly in the urinary tract. It is known for its ability to produce urease, which can lead to the formation of urinary stones and blockages.
P. mirabilis infections are often associated with underlying medical conditions such as diabetes, kidney disease, or urinary catheterization. Symptoms of a P. mirabilis infection may include fever, cloudy or foul-smelling urine, and pain or burning during urination. Treatment typically involves antibiotics that are effective against Gram-negative bacteria, although resistance to certain antibiotics is not uncommon in P. mirabilis isolates.
Proteus vulgaris is a species of Gram-negative, facultatively anaerobic, rod-shaped bacteria that are commonly found in soil, water, and the human digestive tract. They are named after the Greek god Proteus, who could change his shape at will, as these bacteria are known for their ability to undergo various morphological changes.
Proteus vulgaris is a member of the family Enterobacteriaceae and can cause opportunistic infections in humans, particularly in individuals with weakened immune systems or underlying medical conditions. They can cause a variety of infections, including urinary tract infections, wound infections, pneumonia, and bacteremia (bloodstream infections).
Proteus vulgaris is also known for its ability to produce urease, an enzyme that breaks down urea into ammonia and carbon dioxide. This can lead to the formation of urinary stones and contribute to the development of chronic urinary tract infections. Additionally, Proteus vulgaris can form biofilms, which can make it difficult to eradicate the bacteria from infected sites.
In a medical context, identifying Proteus vulgaris is important for determining appropriate antibiotic therapy and managing infections caused by this organism.
Proteus Syndrome is a rare genetic disorder characterized by progressive overgrowth of skin, bones, muscles, and other tissues. It is caused by a mutation in the AKT1 gene, which regulates cell growth and division. The disorder is named after the Greek sea-god Proteus, who could change his shape at will, as people with this condition often have highly variable and asymmetric features.
The symptoms of Proteus Syndrome can vary widely from person to person, but may include:
1. Overgrowth of skin, which can lead to the formation of thickened, rough, or irregular areas of skin (known as "cerebriform" skin) and deep creases or folds.
2. Asymmetric overgrowth of bones, muscles, and other tissues, leading to differences in size and shape between the two sides of the body.
3. The formation of benign tumors (such as lipomas and lymphangiomas) and abnormal blood vessels.
4. Abnormalities of the brain, eyes, and other organs.
5. Increased risk of developing certain types of cancer.
Proteus Syndrome is typically diagnosed based on a combination of clinical features, medical imaging, and genetic testing. There is no cure for the disorder, but treatment is focused on managing symptoms and preventing complications. This may involve surgery to remove tumors or correct bone deformities, physical therapy to improve mobility and strength, and medications to control pain and other symptoms.
"Proteus penneri" is a gram-negative bacterium that is commonly found in the environment, including water and soil. It is a species within the genus Proteus, which are known for their ability to swarm and form spreading colonies on agar media. "Proteus penneri" is closely related to another species, "Proteus mirabilis," and was previously considered to be part of the same species.
"Proteus penneri" can cause a variety of infections in humans, including urinary tract infections, wound infections, and bacteremia (bloodstream infections). It is often resistant to multiple antibiotics, which can make treatment challenging. Proper identification of the organism through laboratory testing is important for guiding appropriate therapy.
It's worth noting that medical definitions can vary depending on the source and context, so it may be helpful to consult a reliable medical or scientific reference for more detailed information.
Urease is an enzyme that catalyzes the hydrolysis of urea into ammonia and carbon dioxide. It is found in various organisms, including bacteria, fungi, and plants. In medicine, urease is often associated with certain bacterial infections, such as those caused by Helicobacter pylori, which can produce large amounts of this enzyme. The presence of urease in these infections can lead to increased ammonia production, contributing to the development of gastritis and peptic ulcers.
Hydroxamic acids are organic compounds containing the functional group -CONHOH. They are derivatives of hydroxylamine, where the hydroxyl group is bound to a carbonyl (C=O) carbon atom. Hydroxamic acids can be found in various natural and synthetic sources and play significant roles in different biological processes.
In medicine and biochemistry, hydroxamic acids are often used as metal-chelating agents or siderophore mimics to treat iron overload disorders like hemochromatosis. They form stable complexes with iron ions, preventing them from participating in harmful reactions that can damage cells and tissues.
Furthermore, hydroxamic acids are also known for their ability to inhibit histone deacetylases (HDACs), enzymes involved in the regulation of gene expression. This property has been exploited in the development of anti-cancer drugs, as HDAC inhibition can lead to cell cycle arrest and apoptosis in cancer cells.
Some examples of hydroxamic acid-based drugs include:
1. Deferasirox (Exjade, Jadenu) - an iron chelator used to treat chronic iron overload in patients with blood disorders like thalassemia and sickle cell disease.
2. Panobinostat (Farydak) - an HDAC inhibitor approved for the treatment of multiple myeloma, a type of blood cancer.
3. Vorinostat (Zolinza) - another HDAC inhibitor used in the treatment of cutaneous T-cell lymphoma, a rare form of skin cancer.
Arachidonate lipoxygenases (ALOXs or ALOXE's) are a group of enzymes that catalyze the dioxygenation of polyunsaturated fatty acids, such as arachidonic acid, to form hydroperoxides. These enzymes play a crucial role in the biosynthesis of various eicosanoids, which are signaling molecules involved in inflammation, immunity, and other physiological processes.
There are several isoforms of ALOXs, including 5-lipoxygenase (5-LOX), 12-lipoxygenase (12-LOX), and 15-lipoxygenase (15-LOX), which differ in their substrate specificity and the position of the hydroperoxide group they introduce into the fatty acid. These enzymes are widely distributed in various tissues, including the lungs, liver, and brain, and have been implicated in a variety of diseases, such as cancer, cardiovascular disease, and neurodegenerative disorders.
Inhibition of ALOXs has been explored as a potential therapeutic strategy for the treatment of these diseases, although the development of selective and safe inhibitors has proven to be challenging.
Benzeneacetamides are a class of organic compounds that consist of a benzene ring, which is a six-carbon cyclic structure with alternating double bonds, linked to an acetamide group. The acetamide group consists of an acetyl functional group (-COCH3) attached to an amide nitrogen (-NH-).
Benzeneacetamides have the general formula C8H9NO, and they can exist in various structural isomers depending on the position of the acetamide group relative to the benzene ring. These compounds are used in the synthesis of pharmaceuticals, dyes, and other chemical products.
In a medical context, some benzeneacetamides have been studied for their potential therapeutic effects. For example, certain derivatives of benzeneacetamide have shown anti-inflammatory, analgesic, and antipyretic properties, making them candidates for the development of new drugs to treat pain and inflammation. However, more research is needed to establish their safety and efficacy in clinical settings.
In the context of medical terminology, tablets refer to pharmaceutical dosage forms that contain various active ingredients. They are often manufactured in a solid, compressed form and can be administered orally. Tablets may come in different shapes, sizes, colors, and flavors, depending on their intended use and the manufacturer's specifications.
Some tablets are designed to disintegrate or dissolve quickly in the mouth, making them easier to swallow, while others are formulated to release their active ingredients slowly over time, allowing for extended drug delivery. These types of tablets are known as sustained-release or controlled-release tablets.
Tablets may contain a single active ingredient or a combination of several ingredients, depending on the intended therapeutic effect. They are typically manufactured using a variety of excipients, such as binders, fillers, and disintegrants, which help to hold the tablet together and ensure that it breaks down properly when ingested.
Overall, tablets are a convenient and widely used dosage form for administering medications, offering patients an easy-to-use and often palatable option for receiving their prescribed treatments.
Methenamine is a medication that is used as a urinary antiseptic. It's a chemical compound that, when ingested and enters the urine, releases formaldehyde, which helps to kill bacteria in the urinary tract. Methenamine is often combined with other medications, such as sodium phosphate or hydroxyzine, to make it more effective.
It's important to note that methenamine is not typically used as a first-line treatment for urinary tract infections (UTIs) and is usually reserved for preventing recurrent UTIs in people who are prone to them. Additionally, methenamine should be taken in adequate amounts and under the guidance of a healthcare professional, as excessive formaldehyde release can cause adverse effects.
 Proteus mirabilis
Proteus mirabilis Proteus Infections: Background, Pathophysiology, Epidemiology
Proteus Infections: Background, Pathophysiology, Epidemiology Pages that link to "Proteus infection medical therapy" - wikidoc
Pages that link to "Proteus infection medical therapy" - wikidoc Serotyping and the Dienes reaction on Proteus mirabilis from hospital infections | Journal of Clinical Pathology
Serotyping and the Dienes reaction on Proteus mirabilis from hospital infections | Journal of Clinical Pathology A small-molecular inhibitor against Proteus mirabilis urease to treat catheter-associated urinary tract infections - Lancaster...
A small-molecular inhibitor against Proteus mirabilis urease to treat catheter-associated urinary tract infections - Lancaster... THE EFFECT OF VERNONIA AMYGDALENA EXTRACT ON PROTEUS INDUCED URINARY TRACT INFECTION IN ADULT WISTAR ALBINO RATS.
THE EFFECT OF VERNONIA AMYGDALENA EXTRACT ON PROTEUS INDUCED URINARY TRACT INFECTION IN ADULT WISTAR ALBINO RATS. Ammonia blood test: MedlinePlus Medical Encyclopedia
Ammonia blood test: MedlinePlus Medical Encyclopedia Advanced Search Results - Public Health Image Library(PHIL)
Advanced Search Results - Public Health Image Library(PHIL) Pathogens | Free Full-Text | Microbial Biofilms in Urinary Tract Infections and Prostatitis: Etiology, Pathogenicity, and...
Pathogens | Free Full-Text | Microbial Biofilms in Urinary Tract Infections and Prostatitis: Etiology, Pathogenicity, and... Treat and Prevent UTIs Without Drugs - Chris Kresser
Treat and Prevent UTIs Without Drugs - Chris Kresser DailyMed - AMPICILLIN injection, powder, for suspension
DailyMed - AMPICILLIN injection, powder, for suspension Guideline for Prevention of
Guideline for Prevention of Tobramycin - Mechanism, Indication, Contraindications, Dosing, Adverse Effect, Interaction, Renal Dose, Hepatic Dose | Drug...
Tobramycin - Mechanism, Indication, Contraindications, Dosing, Adverse Effect, Interaction, Renal Dose, Hepatic Dose | Drug... Ammonia blood test
Ammonia blood test Recurrent Urinary Tract Infections - StatPearls - NCBI Bookshelf
Recurrent Urinary Tract Infections - StatPearls - NCBI Bookshelf Zymox Otic Hydrocortisone Free (8 oz) | On Sale | EntirelyPets
Zymox Otic Hydrocortisone Free (8 oz) | On Sale | EntirelyPets Toxicity Tests | Harvard Catalyst Profiles | Harvard Catalyst
Toxicity Tests | Harvard Catalyst Profiles | Harvard Catalyst Lithostat (Acetohydroxamic Acid Tablets): Uses, Dosage, Side Effects, Interactions, Warning
Lithostat (Acetohydroxamic Acid Tablets): Uses, Dosage, Side Effects, Interactions, Warning Self-organization of swimmers drives long-range fluid transport in bacterial colonies | Nature Communications
Self-organization of swimmers drives long-range fluid transport in bacterial colonies | Nature Communications